ACTIVIDAD Nº ___: QUÉ NOS CUENTAN LOS
HUESOS
INTRODUCCIÓN
Todos los seres vivos tenemos unas características comunes que nos definen
como tales y están incluidas en varios aspectos:
- Composición química, ya que todos están formados por los mismos
bioelementos, los cuales forman las biomoléculas que conforman su
organismo.
- Rutas metabólicas comunes. Las reacciones químicas mediante las que
obtienen la energía que necesita todo organismo para realizar sus
actividades vitales y las rutas metabólicas por las que sintetizan las
biomoléculas son similares.
- La célula es la unidad anatómica y fisiológica que forma y hace posible
el funcionamiento de los seres vivos.
- Código genético común, pues todos los seres vivos mantienen y transmiten
la información de las proteínas que los caracterizan mediante el mismo tipo
de biomolécula: el DNA (ácido desoxirribonucleico).
- Es la uniformidad de estas características lo que ha sugerido que el
origen de la vida sea único y que, con posterioridad, se haya dado el
proceso de diversificación de los seres vivos.
Pues bien, dentro de la gran diversidad de seres vivos que podemos observar
en la actualidad, radica el hecho de que cuantas más semejanzas compartan dos
individuos/ más proximidad filogenética existirá entre ellos. Para establecer
los distintos grupos o taxones que representan el grado de parentesco o
coincidencia morfológica anatómica y funcional, la Filogenia (parte de la
Biología que estudia las relaciones de parentesco) sigue para ello una serie de
procedimientos tales como:
- El estudio de los fósiles, que permite no sólo conocer como eran los
seres vivos en el pasado, sino también establecer series filo gen éticas y,
por tanto, la proximidad evolutiva. Además también se localizan fósiles que
representan por sus características la forma intermedia entre los grupos
posteriormente diferenciados;
- El estudio comparado de la anatomía y la morfología de órganos de seres
vivos ayuda al poner en evidencia la existencia de estructuras homólogas y
análogas. Para que tengan valor filogenético han de presentar un origen
común y una estructura similar: homología, por ejemplo: las extremidades de
los vertebrados, que siguen el mismo diseño estructural (tipo quiridio)
pero con formas diversas ya que su función ha variado con el tiempo:
divergencia adaptativa. En cambio, en las analogías existen similitudes
funcionales: convergencia adaptativa aun siendo de origen diferente y, por
tanto, sin valor filogenético, por ejemplo: alas de insectos y de
vertebrados (pterodáctilo, ave y murciélago).
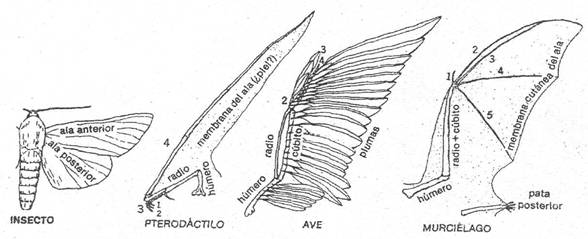
Figura 1: Analogia entre alas de insectos (sin esqueleto
Interno) y de vertebrados (con esqueleto), las cuales tienen una función
similar pero diferente origen. Homología en las alas con el1doesqueleto de
vertebrados, en los terrestres todas se derivan de un patrón común de la
extremidad anterior pero modificada en su morfología. El pterodáctilo (reptil
extinto) presenta un cuarto dedo muy alargado; ave que carece del primero y
quinto, el tercero y el cuarto están parcialmente 1usionados; murciélago con
dedos largos del segundo al quinto. (De Storer et al., General Zoology,
McGraw-Hill Book Company.)
- La embriología (estudio de los organismos desde la fecundación hasta el
nacimiento), ya que en el desarrollo embrionario individual suelen quedar
restos de estados evolutivos anteriores. Lo cual no quiere decir que un
animal pase por todos los estados por los que pasaron sus antepasados de
una forma lineal, sino que los primeros estados embrionarios de todos los
vertebrados son notablemente similares entre sí hasta el momento en que
cada grupo diverge de una manera característica propia. Por ejemplo:
animales de respiración pulmonar en una etapa embrionaria poseen bolsas
faríngeas, que en los peces darán lugar a las hendiduras branquiales pero
en ellos se modifican en diversos sentidos.
- Otros más actuales tales como: pruebas de inmunocompatibilidad,
hibridación de ADN, estudio de las vías metabólicas y de la secuencia de
monómeros de proteínas y ácidos nucleicos (cuanto más próximos
filogenéticamente sean dos seres vivos, mayor será el número de
biomoléculas comunes). Estos últimos procedimientos se complementan con los
anteriores y permiten revisar y perfeccionar las clasificaciones.
DESARROLLO DE LA ACTIVIDAD
Esta actividad se basa en el segundo procedimiento, descrito anteriormente,
y más concretamente en la extremidad tipo quiridio de los vertebrados. Se puede
observar con bastante facilidad que los vertebrados y más concretamente los
tetrápodos poseen la misma estructura en su extremidad anterior, los huesos y
giros o articulaciones de la extremidad son los mismos, existiendo por tanto
una homología y dando a dicha estructura un valor filogenético válido con la
consecuente afirmación de un origen común. Los elementos esqueléticos de dicha
extremidad se disponen según un mismo plan general: en la" parte superior hay
un solo hueso, el húmero, cuya cabeza se articula con la cintura escapular en
la fosa glenoidea. La parte inferior está compuesta por dos huesos paralelos
entre sí, el cúbito y el radio. Por la parte distal de estos huesos hay nueve o
diez huesos carpianos o de la muñeca, cinco metacarpianos más largos en la
mano, y luego unas hileras de pequeñas falanges en los dedos.
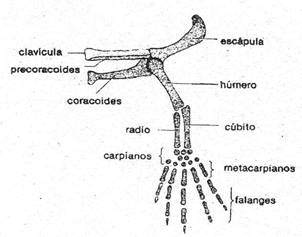
Figura 2: Esqueleto de la cintura y extremidad torácicas de
los vertebrados. La clavícula presenta un punteado más claro para indicar que
es un hueso membranoso.
También se observa claramente que la disposición, tamaño, grosor... de los
huesos es distinto según el grupo de tetrápodos de que se trate, esto es así ya
que cada ser vivo está adaptado al medio en el que vive y no es sino a partir
de lo que ya se 'tiene que se puede cambiar, esto quiere decir que a partir de
una misma estructura y gracias a la variabilidad gen ética (posibilidad de
cambio) ya la interacción con el medioambiente cambiante, a lo largo de la
evolución se han ido adaptando a los diferentes hábitats dando lugar a
divergencias adaptativas. La mayoría de las especializaciones consisten en la
reducción o fusionamiento de diversas partes del esqueleto pentadáctilo
primitivo.
- Están adaptadas al vuelo las extremidades del murciélago y del ave, pero
de forma muy diferente: en él los huesos se han convertido en una
estructura en la que se insertan las plumas, el húmero es grande y ancho y
ofrece una gran superficie para que se inserte la musculatura pectoral, el
radio y la ulna también son grandes y la mano está formada por tres dedos,
de los que sólo el primero puede realizar movimientos independientes. En el
murciélago o quiróptero, que significa " ala en la mano" , el antebrazo
(radio y cúbito, el cual suele estar muy reducido existiendo en algunos'
casos sólo su porción proximal estando la parte distal fusionada con el
radio) y cuatro dedos de la mano están extraordinariamente alargados a modo
de varillas que sostienen un pliegue cutáneo o patagio, muy vascularizado,
que se extiende también por los lados del cuerpo hasta llegar a los
miembros posteriores, dejando los pies libres, y la cola; generalmente el
dedo pulgar queda libre de membrana y tiene uña.
- Como adaptación a la natación, las extremidades anteriores de la ballena
están transformadas en aletas actuando como remos rígidos que ayudan al
equilibrio ya la dirección al acortarse y ensancharse los huesos proximales
(húmero, radio y cúbito) y al multiplicarse las falanges de los dedos
centrales.
- El lagarto es un ejemplo de reptil en los que la mayoría del peso (en
cuanto a la extremidad anterior) recae sobre el radio con lo que el cúbito
ha sufrido una regresión que le ha hecho disminuir de tamaño. (Esta es
también la razón por la que los reptiles no puedan realizar los movimientos
de pronación, rotación de la mano de fuera a dentro, y supinación, rotación
de la mano de dentro a fuera). El metacarpo y las falanges son
alargados.
- Adaptado a una marcha rápida en la que el contacto entre el suelo y la
extremidad se da únicamente por los dedos: digitígrado, tenemos como
ejemplo al gato, el cual además presenta un alargamiento de la mitad
inferior de las extremidades, lo que le confiere una carrera veloz y unos
lances potentes y ágiles. Posee cinco dedos, el primero muy alto no deja
huella en el suelo.
- La configuración de la extremidad anterior del humano le proporciona una
gran libertad de movimiento: la mano está adaptada a la prensión gracias a
la existencia del pulgar oponible, la muñeca posee gran capacidad de
movimiento, los huesos proximales, alargados y. bien desarrollados permiten
la inserción de los músculos realizando así una gran cantidad de
movimientos ej.: movimientos de pronación y supinación realizados por el
antebrazo (donde intervienen radio y cúbito).
- La rana es un anfibio que posee una extremidad anterior robusta, cuyo
húmero y falanges (la primera de las cuales. está atrofiada) son muy
alargadas en relación al radio y al cúbito, los cuales tienden a
fusionarse. Complementa a la extremidad posterior en la natación ya que
ésta es el verdadero órgano de propulsión de este animal.
La extremidad tipo quiridio es originaria de los tetrápodos y tiene su
homología en una aleta (los peces también son vertebrados): hay un grupo, el de
loS Osteolepiformes (pez crosopterigio ripidistio fósil), y más en concreto
Eusthenopteron, que se admite como muy cercano por sus caracteres a los
primeros tetrápodos, cuya aleta lobulada es monobasal (un solo elemento se
articula con la cintura) y dicotómica (cada elemento se articula distalmente
siempre con dos), lo cual también se puede observar, prescindiendo de las
falanges, en un quiridio de anfibio primitivo. Se cree que el primer hueso de
la base de la aleta es equivalente al húmero y el segundo al cúbito; el primer
radial a dicho cúbito es comparable al radio y el resto de los radiales se
considera que se han convertido en los huesos carpianos, mientras que los
metacarpianos y falanges han surgido como nuevas excrecencias al borde de la
porción muscular carnosa de la aleta. El mismo Eusthenopteron no se considera
como precursor directo de los tetrápodos, sino que éstos surgirían de algún
otro ripidistio (se han encontrado fósiles de anfibios primitivos junto con los
de los ripidistios, en los mismos depósitos geológicos).
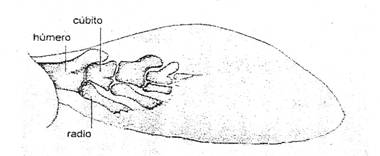
Figura 3: Esqueleto de la aleta pectoral izquierda del pez
crosopterigio Eusthenopteron
Se pensaba que el quiridio primitivo era pentadáctilo, pero hay anfibios
primitivos con hiperfalangia; o sea que con el quiridio se pierden las aletas,
pero al seguir en medio acuático aparece una hiperfalangia que se reduce al
pasar al medio terrestre, con lo que la pentadactilia es una adaptación a dicho
medio y no una adaptación de los tetrápodos. Además en ambas extremidades se
producen giros distintos: codo y rodilla y tobillo y muñeca que favorecen el
desplazamiento y se encuentran más pegados al cuerpo presentando una locomoción
más eficaz.
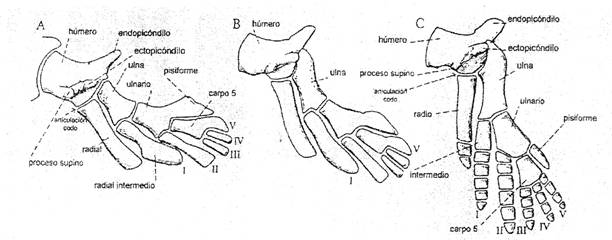
Figura 4: Diagrama que ilustra los principales cambios en la
transformación de la aleta pectoral osteolipiforme en extremidad tetrápoda. A,
estado Eusthenopteron. B, estado intermedio. C, estado tetrápodo primitivo
Ha de quedar claro que si aceptamos la teoría del origen común de las
especies, todas están igual de "'evolucionadas" y la diferencia radica en el
grado de complejidad que poseen dichas especies. No hay un ser vivo más
evolucionado que otro, sino que con el transcurso de la evolución unos han ido
surgiendo a partir de otros anteriores y otros muchos han ido desapareciendo al
ir variando las condiciones del medio y que los individuos que existen en un
momento concreto lo hacen gracias a que poseen la capacidad de adaptarse al
medio en el que viven (poseen las características que le permiten sobrevivir en
un cierto ambiente).
Daniel Tomás. IES Abastos, Valencia.
VERSIÓN
PDF PARA IMPRIMIR
VOLVER AL ÍNDICE DE PRÁCTICAS